预期的感觉信息从肌肉收缩抑制不良被认为是高疲劳的基础。这种与任务相关的感觉衰减减弱反映在静息状态连接中。在这里,我们检验了“脑电图(EEG)衍生的β带体感网络功能连接的改变是中风后疲劳的一个特征”的假设。
在无抑郁、最小损伤的中风幸存者(n=29)中位病程为5年,使用64通道脑电图测量静息状态神经元活动。在beta (13-30 Hz)频率范围内,通过小世界指数(SW)计算基于图论的网络分析功能连通性度量,重点关注左右运动(Brodmann区4、6、8、9、24和32)和感觉(Brodmann区1、2、3、5、7、40和43)网络。使用疲劳严重程度量表- FSS(卒中)测量疲劳,得分> 4,定义为高度疲劳。
结果证实了工作假设,与低疲劳相比,高疲劳中风幸存者在体感觉网络中表现出更高的小世界。
体感网络中高水平的小世界表明对躯体感觉输入的处理发生了改变。这种加工过程的改变可以解释疲劳的感觉衰减模型中的高努力知觉。
中风是神经血管功能不全的结果,以疲劳为主要症状。中风的严重程度并不能解释报告的疲劳程度(Kutlubaev et al. 2012;van der Werf et al. 1998)。之前,我们提出了疲劳的感官衰减假设,基于预期感官信息的抑制不良- (Kuppuswamy 2017,2022)。肌肉感觉传入抑制不良导致对简单任务分配高努力,这解释了卒中后疲劳的很大一部分(Doncker等人,2020a, b)。在视觉和听觉感知中,干扰物抑制不良解释了卒中后疲劳(Kuppuswamy等人,2022;Doncker and Kuppuswamy 2022)。虽然其他大脑区域/网络也与疲劳有关,如顶叶、前额叶和皮层下网络(Jaeger等人,2019;Finke et al. 2015;Cotter et al. 2021),这里我们特别关注与PSF有关的体感和运动网络;为了验证感觉衰减假说的预测,即疲劳是由体感觉网络的功能障碍引起的。
从神经网络功能的角度来看,行为反映了网络的结构和功能变化,这些变化在静息状态下持续存在(Graziadio et al. 2010)。静止的神经元网络表达了跟踪其执行所需行为的能力的特征(Kim和Kang 2018;Wahlheim et al. 2022;Doucet et al. 2012;李艳等。2022a, b;Liu et al. 2022)。静止网络的这些特征显示出反映慢性症状的变化(Porcaro et al. 2019)。卒中后疲劳(PSF)和其他疲劳显著的疾病,神经生理(Kuppuswamy et al. 2015 5a, b, c;ondobaka et al. 2021;De Doncker et al. 2021;Liepert et al. 2005;Morgante et al. 2011;Russo et al. 2017)和行为学(Kuppuswamy et al. 2015a, b, c, 2016;De Doncker等。2020a;Doncker等人(2020b)的研究结果支持静息状态的改变,特别是感觉网络活动。
以特定频率放电并通过同步放电相互交流的神经元集合组成了一个神经元网络。为了了解网络的活动,使用功能连接方法映射各个节点之间的同步性强度(Bullmore和Sporns 2009)。功能连通性被定义为不同神经元群和区域之间的时间相关性或依赖性(Fingelkurts et al. 2005;Rubinov and Sporns 2010)。这种时间相关性发生在不同的频段,低频与觉醒有关,中频与感觉运动活动有关,高频代表高阶函数,如错误检测和学习。由于PSF被认为是感觉运动控制的一个问题,特别是在处理传入的肌肉相关感觉信息时产生的,我们预计疲劳相关的β频带频率调制。
在这里,我们研究了体感觉网络的功能障碍是否导致了PSF,正如休息时感觉和运动网络中β带神经元活动的变化所证明的那样。
本研究已获伦敦布罗姆利研究伦理委员会批准(REC参考编号:16/LO/0714)。中风幸存者被招募并在英国伦敦神经病学研究所接受测试。
所有中风幸存者在研究前都根据以下标准进行筛查:首次缺血性或出血性中风;在研究前至少3个月发生中风;无其他神经系统疾病的临床诊断;中风后身体恢复良好,定义为受影响手的握力和手灵巧度至少为未受影响手的60%,分别使用手持式测力仪和九孔钉测试(NHPT)进行评估;未服用抗抑郁药或其他对中枢神经系统有直接影响的药物;使用医院焦虑和抑郁量表(HADS)(Snaith 2003)评估抑郁评分≤11的非临床抑郁患者。
29名中风幸存者参加了这项研究(表1),并根据赫尔辛基宣言提供了书面知情同意书。由于缺乏试验数据,没有进行正式的样本量计算。然而,在之前的研究中,每组只有10名受试者在静息状态功能连接的测量中可以观察到差异(Nordin et al. 2016)。
采用疲劳严重程度量表FSS-7对性状疲劳进行量化。平均得分为1表示没有疲劳,而平均得分为7表示极度疲劳(Krupp et al. 1989)。高疲劳定义为FSS-7 > 4 (Valko et al. 2008)。
对照组:本研究没有招募健康的人作为对照组,特别是因为大脑的状态很可能在受伤或患病后发生变化(Kuppuswamy 2023),因此理想的对照组将是疾病内对照组。卒中低(无)疲劳组作为本研究的对照组。
全头皮脑电图(EEG)数据记录使用64通道系统,ActiCap, Herrsching,德国,和BrainAmp,休息时,眼睛睁开并聚焦于固定交叉。录音时长为7分钟。64个电极按照10-20个国际EEG电极阵列放置在帽上。在线录音时,分别以通道FCz和通道AFz作为基准和地。在整个记录过程中,阻抗保持在10 kΩ以下。EEG信号以1khz采样,并使用BrainVision Recorder软件(BrainVision Recorder, Version 1.21.0102 Brain Products GmbH, Gilching, Germany)在线可视化。
结合EEGLAB (Delorme and Makeig 2004)和自定义Matlab脚本进行EEG分析。EEG数据下采样至250 Hz,然后使用有限脉冲响应滤波器从0.1 Hz带通滤波至47 Hz。使用自动化程序识别和去除噪声通道。随后,将脑电数据分割为2个second epoch,对包含噪声数据的epoch进行识别:计算所有脑电通道的平均活动,并将阈值设置为平均活动标准差的±2倍。包含超过阈值的活动的时代被标记并随后被删除。这样总共剩下160(±15)个两秒的周期。为了从EEG数据中识别和去除眼球运动和眨眼伪影,在EEGLAB中实现了独立分量分析(ICA)。ICA是一种盲源分解算法,可以从多通道数据中分离出统计独立的源(Jung et al. 2000)。随后对这些组件进行目视检查,并去除那些包含眼球运动或眨眼伪影的组件。然后将先前删除的通道内插回数据集,最后,将脑电图数据与所有头皮电极的大平均值重新引用。
功能连通性分析。EEG连通性分析使用精确低分辨率电磁断层扫描(eLORETA)软件进行(苏黎世精神病学大学关键研究所;http://www.uzh.ch/keyinst/NewLORETA/LORETA01.htm)。eLORETA算法是一种行之有效的EEG信号线性逆解(Pascual-Marqui 2002)。
全脑源重建后,使用eLORETA软件计算四个脑区的连通性,根据Broadmann区(BAs)将其分为左右脑半球的运动和感觉网络。每个BA是一个感兴趣的区域(ROI)。形成左右半球运动网络的ba包括BA4、6、8、9、24和32,形成左右半球感觉网络的ba包括BA1、2、3、5、7、40和43。
在eLORETA中计算每个网络中所有BAs的电流密度时间序列,并使用滞后线性相关(LagR)算法估计功能连通性,不受每个网络中的体积电导和低空间分辨率的影响(Pascual-Marqui 2007)。计算beta (13-30 Hz)频带频率的滞后线性相干性。
图分析。网络是现实世界复杂系统的数学表示,由节点对之间的节点(顶点)和链接(边)的集合定义。大规模大脑网络中的节点代表大脑区域,而链接代表解剖或功能连接。理想情况下,淋巴结应该代表具有连贯的解剖或功能连接模式的大脑区域。提取各频段所有对roi之间的连通性参数为方阵W,其维数等于roi的个数。矩阵W内的每一行和每一列表示节点,矩阵内的值表示每对节点之间的连接强度。
一旦构建了感兴趣的网络,就可以使用Brain Connectivity Toolbox (Rubinov and Sporns 2010)计算总结网络隔离和集成方面的图论核心度量。聚类系数(C)是指网络元素形成独立集群的程度,用聚类系数(C)来衡量。整合是指网络相互连接和交换信息的能力,用参数特征路径长度(L)来衡量。聚类系数和特征路径长度分别代表网络在局部连通性和全局连通性方面的效率。加权聚类系数(Cw)和加权特征路径长度(Lw)作为网络分离和整合的度量,计算如下:
其中,Cbrain和Lbrain是由每个参与者的连接矩阵导出的聚类系数和特征路径长度。Crandom和Lrandom是100个与原始网络具有相同基本特征的代理随机网络的聚类系数和特征路径长度的平均值,这些网络是利用原始网络的边权随机重新布线而得到的。随机网络保留了与原始网络相同的基本特征,如节点数量和边缘权重。随机网络由Brain Connectivity Toolbox29构建。因此,网络小世界度(Sw)的度量被定义为Cw与Lw之间的比率;本地连通性与网络的全球整合之间的比率。
当Sw的值约为1时,网络被称为具有“小世界属性”,这意味着节点之间的高水平局部聚类和全局连接所有网络节点的适当路径的良好组合(大系统的所有节点通过相对较少的中间步骤连接)。Sw值大于1表明节点之间的局部聚类程度高,并且有许多短路径全局连接网络的所有节点,而Sw值小于1表明本地连通性差,连接发育不良。
所有统计分析均使用R (RStudio Version 1.2.5033)进行。Spearman秩相关确定了特质疲劳(FSS-7)与人口统计学变量(年龄、握力、NHPT、HADS -抑郁、HADS -焦虑和中风后时间)之间的关联。Wilcoxon秩和检验确定了特质疲劳(FSS-7评分)和分类测量(性别、受影响半球和中风类型)之间的关联。由于各组血管面积太少,未分析血管面积对FSS的影响。Shapiro-Wilk正态性检验评估因变量分布,Levene检验评估方差齐性。
在正态分布变量中,采用三向混合方差分析来评估疲劳(受试者因素之间:StrokeLow, StrokeHigh)和性别(受试者因素之间:男性,女性)对beta频带(因变量)内小世界的影响,涉及两个网络(感觉和运动)和两个半球(左半球和右半球)。温室-盖瑟调整修正了任何与球度的偏差。采用Bonferroni调整进行事后两两比较(t检验)以确定主要影响。
摘要
介绍
方法
结果
讨论
结论
数据可用性
参考文献
致谢
作者信息
道德声明
电
nic补充资料
搜索
导航
#####
29名中风幸存者完成了这项研究(11名女性和18名男性)。FSS-7评分中位数女性为5.29 (IQR=2.57),男性为2.50 (IQR=2.46)。经Wilcoxon检验,FSS-7评分差异无统计学意义(p=0.05,效应量=0.37)。特质疲劳(FSS-7)与所有连续人口学测量之间的Spearman秩相关显示,特质疲劳与HADS-Depression之间存在显著正相关(Spearman ρ=0.41, p=0.03),而其他变量与特质疲劳无关(年龄:Spearman ρ=0.02, p=0.91;握力:Spearman ρ=-0.23, p=0.24;NHPT: Spearman ρ=-0.25, p=0.19;hads -焦虑:Spearman ρ=0.35, p=0.06)。
表1此表提供了卒中幸存者队列的人口学和病变信息
在这项研究中,没有任何中风幸存者被证实有核磁共振损伤。在所有中风幸存者中评估特质疲劳(FSS-7)与中风临床特征之间的关系。右半球卒中患者FSS-7得分中位数为4.43 (IQR=3.00),左半球卒中患者FSS-7得分中位数为2.71 (IQR=3.75) (Wilcoxon检验:p=0.50,效应量r=0.13)。缺血性卒中患者FSS-7评分中位数为2.93 (IQR=3.46),出血性卒中患者FSS-7评分中位数为3.86 (IQR=2.29) (Wilcoxon检验:p=0.51,效应量r=0.13)。关于受影响的血管区域,由于无法检索到临床记录,因此缺少来自5名中风幸存者的数据。中动脉病变组FSS-7评分中位数为3.36 (IQR=3.25),主成分病变组FSS-7评分中位数为3.07 (IQR=2.07),脑干/小脑病变组FSS-7评分中位数为4.21 (IQR=2.68) (Kruskal-Wallis检验:p=0.57,效应大小η2=-0.04)。FSS-7与参与者参加研究的中风后时间之间的spearman等级相关性显示无显著相关性(spearman ρ=0.08, p=0.67)。在目前的中风幸存者队列中,由于数据不准确,很难对中风类型和血管范围对FSS-7的影响做出有意义的解释。
在两个网络(感觉/运动)和半球(左/右)中有两个极端的异常值(都在右半球的运动网络中)。排除异常值后,数据符合正态分布(p > 0.05),方差均匀(p > 0.05)。三因素方差分析显示,网络类型对beta频段小世界度的影响主要为F(1,23)=15.59, p < 0.01, η2=0.13,网络类型与疲劳程度之间存在显著交互作用(F(1,23)=16.79, p < 0.01, η2=0.14)。网络类型、疲劳程度和性别之间也存在显著的双向交互作用(F(1,23)=5.09, p=0.03, η2=0.05),但性别与网络类型之间不存在显著的双向交互作用(F(1,23)=0.65, p=0.43, η2=0.006),性别与疲劳之间不存在显著的双向交互作用(F(1,23)=1.05, p=0.32, η2=0.009)。事后多组两两比较显示右半球感觉网络中StrokeLow组和StrokeHigh组在beta频段内的小世界(图1)存在显著差异(p=0.01),这是由女性参与者的差异驱动的(p=0.0127)。StrokeLow和StrokeHigh在左右脑运动网络上也存在显著差异(p=0.02和p=0.03),其中女性在右脑运动网络上的差异最大(p=0.0138)。
图1
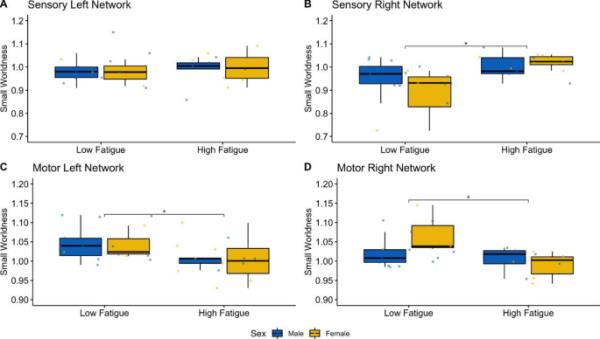
感觉和运动网络在β频带频率上的功能连通性。在男性和女性的两个疲劳组(蓝色为StrokeMale,黄色为StrokeFemale)中,小世界的值通过左(A)和右(B)半球的感觉网络以及左(C)和右(D)半球的运动网络的箱形图显示出来。疲劳组间差异以星号表示(*=p < 0.05)。在右侧的运动和感觉网络中,这种差异是由女性驱动的
在29例中风患者中,我们发现在β频带频率(13-30 Hz)中,高水平的特质性疲劳患者在感觉网络中表现出明显较高的小世界水平,而在运动网络中表现出明显较低的小世界水平。中风的临床特征与特质性疲劳之间没有关联,这证实了先前的发现(Kutlubaev et al. 2012)。根据小世界的研究,脑功能连接同时调和了功能整合和功能分离的对立需求。小世界指数反映了功能专门化(隔离)模块与大量模块间(集成)链接之间的平衡。在这里,我们观察到疲劳与感觉和运动网络中小世界组织的改变是平行的。
在感觉网络中观察到的变化是侧向的。一个微不足道的解释可能是,小数字导致一方达到重要意义,而另一方却没有。如果观察到的侧化不是由于数量有限,那么右侧感觉网络中的“超连接”与最近的发现一致,即在中风后高度疲劳的患者中,感觉运动网络向右半球主导转变(ondobaka et al. 2021)。健康的大脑表现出左半球优势(Netz et al. 1995;Giovannelli et al. 2009),在包括抑郁症在内的几种精神疾病中也观察到向右主导的转变(Lefaucheur et al. 2008)。第三种可能的重要解释可能是低疲劳组左半球卒中数量的微小差异,然而,这是不太可能的,因为先前的研究表明,无论卒中的一侧如何,半球优势都会发生变化(ondobaka et al. 2021)。
β带活动通常被称为感觉运动“空转”节律,在所有皮层和皮层下运动区休息时都可以看到。运动使β带振荡不同步,从而导致β频率是感觉运动区域休息的节奏的想法。最近的研究表明,β带活动可能不仅仅反映缺乏运动,而是维持感觉运动现状的一个指标(Engel和Fries 2010)。在静息β带活动自发增强期间,运动比静息β带活动较低时慢(Gilbertson et al. 2005)。根据这些发现,目前的结果表明,那些高度疲劳的人很可能动作迟缓。在脑卒中后疲劳中,虽然反应时间没有差异,但运动速度减慢(Kuppuswamy et al. 2015a, b, c),这可能是因为存在改变感觉运动现状的阻力,正如本研究中显示的增强的β节律小世界。
众所周知,疲劳与运动和认知缺陷的关系非常有限(Ingles et al. 1999;Winward et al. 2009)。然而,尽管有良好的功能能力,行为灵活性差的标志与高度疲劳有关(De Doncker等人。2021;Morgante et al. 2011)。增强的感觉和减少的运动网络功能连接,寻求维持一种感觉状态,从而使新的感觉状态不那么理想,进一步支持了行为灵活性差的观点,支持高度疲劳。
在这项研究中,我们提供的证据表明,在改变体感觉加工,这可能表明一个可能的机制,驱动疲劳是不良的体感觉加工。在脑卒中后疲劳和其他神经系统疾病(如多发性硬化症)中,有几份报告显示静息状态连通性改变(ondobaka等人,2021;Woodward et al. 2019;Bisecco et al. 2018;Jaeger et al. 2019;Stefancin et al. 2019;Cotter et al. 2021)。在中风中,顶叶连接不足和额叶连接过度(Cotter etal . 2021)以及半球间连接平衡逆转(ondobaka etal . 2021)与疲劳表现有关。在MS中,默认模式网络的变化(Jaeger等人,2019)和涉及运动、感觉和动机的纹状体回路的参与(Bisecco等人,2018)都与疲劳的发展有关。虽然有几个大脑区域作为疲劳的核心区域与这两种疾病有关,但很少有研究对静息状态活动进行假设驱动的分析。在本研究中,我们假设对躯体感觉输入的注意不像正常情况那样受到抑制[表明在肌肉收缩过程中感知到的高度努力(Doncker等人,2020a, b)],这将在静息状态下反映在感觉和运动网络中。感觉网络的超连通性通常被视为维持感觉状态现状倾向的标志,而由此导致的运动网络的低连通性表明M1兴奋性较低(Kuppuswamy等人,2015a, b, c),支持了体感衰减不良支撑高PSF的前提。总之,体感网络中小世界性的增加表明了维持现状的倾向,即休息;不作为(休息)倾向的增加转化为发起行动所需的努力的增加;增加努力会导致疲劳。
与低疲劳组相比,高疲劳组的女性人数不成比例地高。先前的研究报告称,女性中风幸存者的疲劳发生率更高(Cumming et al. 2016),性别显著影响静息状态连通性的测量(Stumme et al. 2020)。虽然疲劳发生率的差异先前归因于诸如受社会文化因素影响的报告偏差等因素,但本研究中发现的高度疲劳的男性和女性之间静息状态连通性的差异,为PSF发病率差异的生物学基础提供了可能性。性别对静息状态连通性的影响不太可能是这里看到的差异的驱动因素,因为感觉运动网络不太可能受到性别的影响(L. Li et al. 2022),不像其他大脑网络。虽然进一步推测男性和女性驱动疲劳的生物学机制差异超出了本文的范围,但未来的疲劳机制研究必须考虑性别可能是解释研究结果的混杂因素,并直接研究男性和女性在PSF中的差异。
局限性:虽然本研究的假设驱动方法是一种优势,但它也可能是一种局限性。大脑中没有一个神经网络是孤立运作的,其他网络对大脑感觉运动区域的影响也可能受到本研究中发现的差异的影响,这需要进一步研究。这项研究的参与者人数相对较少,这进一步突出了研究中两性之间的差异。病变位置不影响PSF的发生率或严重程度,然而,为了明确排除病变位置对与疲劳相关的神经网络活动的影响,每种病变类型都需要更大的数据。
慢性疲劳是几种已确定的长期疾病的特征。然而,没有一个原则性的框架来理解疲劳的机制。在本文中,我们证明感觉运动网络活动的改变符合慢性中风幸存者疲劳的感觉衰减模型的预测。这是一个有希望的框架,可以解释其他神经系统疾病与长期疲劳的连接改变,未来的工作必须集中在探索慢性疲劳的感觉衰减。
以下是电子补充材料的链接。
下载原文档:https://link.springer.com/content/pdf/10.1007/s10548-023-00975-8.pdf
为您推荐:
- 总统:我相信,我们将继续共同努力,进一步扩大阿塞拜疆与阿塞拜疆的关系 2025-07-21
- |天空广播倒数10部最伟大的天气电影 2025-07-21
- 考特妮·卡戴珊向已故父亲罗伯特·卡戴珊致敬 2025-07-21
- 拉什达·琼斯与“最喜欢的双鱼座伙伴”庆祝48岁生日 2025-07-21
- 南非骑师沃伦·肯尼迪在一天内赢得7场比赛,包括在普凯科赫的第一组 2025-07-21
- Daryl Hall tour 2023:在哪里买票,时间表,价格 2025-07-21


